Bartosz Szymański
Wydział Biologii UW
Koło Naukowe Biologii Molekularnej UW
Aminokwasy to związki organiczne zawierające w swojej strukturze grupę kwasową (zazwyczaj karboksylową (-COOH)) i jednocześnie grupę aminową (-NR2 gdzie R = H lub podstawnik węglowy aromatyczny bądź alifatyczny). Grupa karboksylowa zawiera kwasowy atom wodoru, który w wyniku dysocjacji odchodzi od cząsteczki aminokwasu w postaci kationu H+ (proton), pozostawiając stabilizowany przez delokalizację elektronów anion karboksylanu. Tak się składa, że w obrębie tej samej cząsteczki znajduje się aktywny „wychwytywacz” protonów, jakim jest grupa aminowa, ze względu na obecność wolnej pary elektronowej atomu azotu.
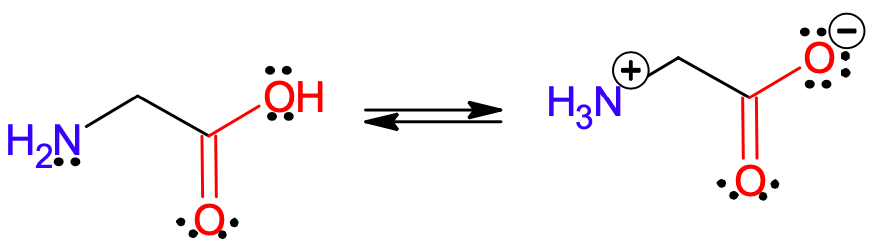
Ryc. 1. Transfer protonu w aminokwasach na przykładzie glicyny
Produkt takiego transferu protonu jest nazywany jonem obojnaczym, ze względu na obecność w tym indywiduum dwóch separowanych ładunków o przeciwnych znakach. Inna nazwa jonu obojnaczego to z języka niemieckiego zwitterion. Ze względu na obecność grupy karboksylowej warunkującej właściwości kwasowe i grupy aminowej warunkującej właściwości zasadowe, aminokwasy reagują zarówno z kwasami, jak i z zasadami. Dla każdego aminokwasu można określić takie pH, w którym statystycznie wszystkie cząsteczki nie posiadają nadmiarowego ładunku elektrycznego – czyli występują w postaci jonów obojnaczych. Jest to tak zwany punkt izoelektryczny, którego wartość jest charakterystyczna dla każdego aminokwasu i może posłużyć do rozdziału mieszaniny aminokwasów. Jeśli obniżymy pH, czyli zwiększymy stężenie jonów H+, to aminokwas będący wcześniej w punkcie izoelektrycznym zostanie sprotonowany na grupie karboksylowej. Z kolei gdy podwyższymy pH, dodając mocnej zasady, to aminokwas będący w punkcie izolelektrycznym zostanie zdeprotonowany na grupie aminowej.
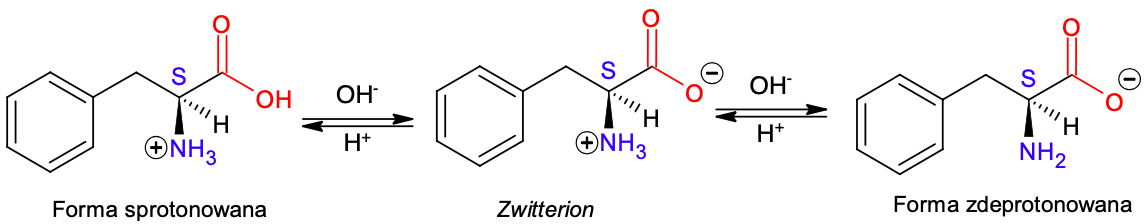
Ryc. 2. Postać aminokwasu w zależności od pH na przykładzie L-fenyloalaniny
Interesująca jest różnorodność punktów izoelektrycznych dla poszczególnych aminokwasów – od 2,77 dla kwasu asparaginowego do 10,76 dla argininy. Ta rozbieżność wynika z różnej kwasowości i zasadowości aminokwasów. Aminokwasy kwasowe są bardziej intuicyjne do określenia – są to wszystkie aminokwasy zawierające nadmiar grup karboksylowych w stosunku do grup aminowych. I tak najbardziej znane aminokwasy kwasowe to kwas glutaminowy i kwas asparaginowy. Natomiast ciekawiej sytuacja wygląda wśród aminokwasów zasadowych – tutaj nie tylko nadmiar grup aminowych powoduje właściwości zasadowe, zamiast grup aminowych możemy mieć na przykład pierścień imidazolu (cykliczna amidyna – przykładem aminokwasu z pierścieniem imidazolu jest histydyna), lub fragment guaniny obecny w argininie.
Oprócz wymienionych dotąd histydyny i argininy, wśród aminokwasów zasadowych znajdują się między innymi lizyna i ornityna (ta ostatnia nie należy do aminokwasów białkowych). Aminokwasy białkowe to takie, które wchodzą w strukturę białek. Ponieważ aminokwasy są w organizmach wprowadzane do białek na podstawie trójkowego kodu genetycznego składającego się z oznaczeń zasad azotowych obecnych w RNA, to logiczne jest, że skoro w RNA mamy tylko cztery różne zasady azotowe, to maksymalna możliwa liczba kombinacji ich różnych trójek wynosi 43 = 64. Jest to maksymalna możliwa liczba aminokwasów białkowych. Okazuje się, że w rzeczywistości ich liczba jest mniejsza, ponieważ kod genetyczny jest zdegenerowany – to znaczy, że dany aminokwas może być kodowany przez więcej niż jedną trójkę zasad azotowych. Finalnie dwadzieścia aminokwasów zyskało miano aminokwasów białkowych kanonicznych. Ponadto wyróżnia się jeszcze dwa niekanoniczne aminokwasy białkowe, czyli selenocysteinę (analog cysteiny) i pirolizynę. Ich niekanoniczność wynika stąd, że są one kodowane przez trójki zasad azotowych normalnie oznaczających kodon STOP. Jednak dla niektórych organizmów kod genetyczny jest minimalnie różny, więc możliwe jest kodowanie nadprogramowych aminokwasów.
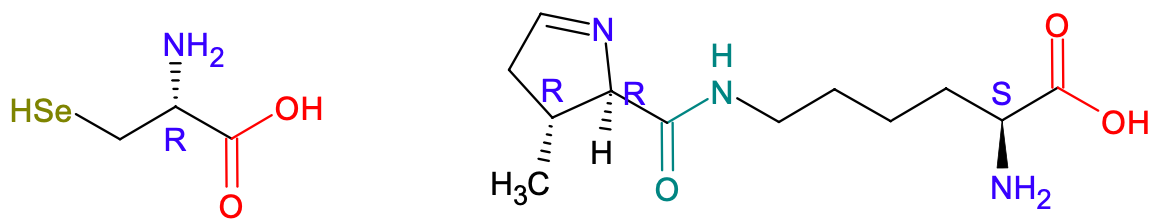
Ryc. 3. Cząsteczki L-selenocysteiny (z lewej) i L-pirolizyny. Selenocysteina różni się od cysteiny tylko atomem selenu, który został wprowadzony zamiast atomu siarki. Reaktywność związków siarki i selenu jest bardzo podobna. Natomiast pirolizyna na pierwszy rzut oka wygląda jak dipeptyd - i całkiem słusznie, na niebiesko-zielono zaznaczyłem wiązanie amidowe.
Najważniejszą z biologicznego punktu widzenia reakcją, której ulegają aminokwasy, jest tworzenie peptydów. Wiązanie C=O posiada moment dipolowy skierowany w kierunku atomu tlenu, przez co na atomie węgla znajduje się cząstkowy ładunek dodatni. Do tego dodatniego ładunku zbliżają się elektrony z wolnej pary elektronowej atomu azotu z grupy aminowej. Należy jednak zachować ostrożność podczas rozważania tworzenia wiązania peptydowego! Oto wiemy, że grupa aminowa w aminokwasie jest zasadowa, a grupa karboksylowa ma właściwości kwasowe – dlaczego w wyniku ich reakcji miałoby powstać wiązanie peptydowe, a nie na przykład sól, jak w typowej reakcji kwasu i zasady? Rozwiązaniem tej zagadki jest synteza aktywnych „estrów” na etapie wstawiania kolejnego aminokwasu w rybosomie – konkretnie są to addukty aminokwasu i tRNA, znane jako aminoacylo-tRNA, dzięki czemu produktem jest nie sól, a peptyd.
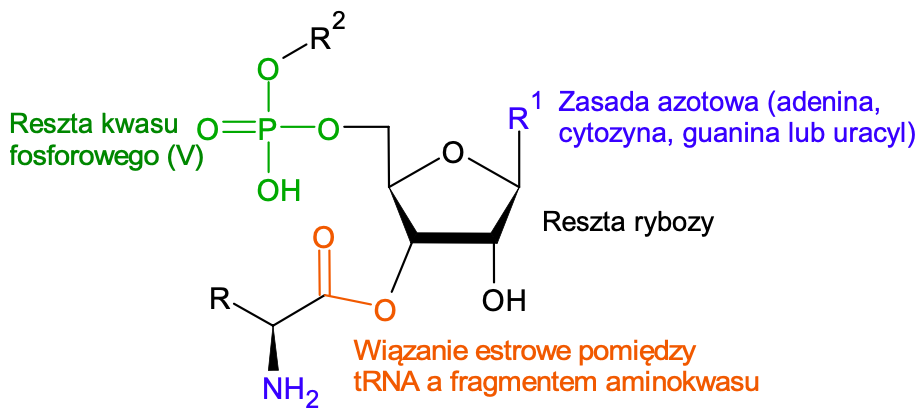
Ryc. 4. Schemat budowy aminoacylo-tRNA: R oznacza łańcuch boczny aminokwasu, pozostałe istotne elementy w aminoacylo-tRNA zostały podpisane. Przydatność tworzenia wiązania estrowego jest nieoceniona - w reakcji estru i grupy aminowej nie może powstać sól, ponieważ grupa estrowa nie wykazuje właściwości kwasowych.
W kolejnym etapie do utworzonego estru przyłącza się kolejny aminokwas (grupą aminową) i powstaje wiązanie peptydowe, oraz odtwarza się cząsteczka tRNA. Można więc powiedzieć, że tRNA jest katalizatorem reakcji tworzenia wiązania peptydowego. Synteza laboratoryjna peptydów czerpie z dokonań ewolucji – chemicy organicy stosują podobną metodę syntezy peptydów, opartą na tworzeniu aktywnych estrów.
Aminokwasy posiadają łańcuch boczny; szczególnie w aminokwasach białkowych powszechne jest nazywanie fragmentu cząsteczki łańcuchem bocznym. Dla aminokwasów białkowych łańcuchem bocznym nazywamy tę część cząsteczki, która jest specyficzna dla danego aminokwasu. Często nieco się ją deprecjonuje, skracając ją we wzorze aminokwasu do „R” – podczas gdy wygląda na to, że jest to fragment cząsteczki niezwykle istotny dla właściwości przyszłych białek – nawet tych właściwości obserwowalnych w makroskali, jak giętkość włosów; tutaj duże znaczenie ma tworzenie mostków disiarczkowych pomiędzy łańcuchami bocznymi cysteiny.
Warto jeszcze w tym miejscu przedstawić zwyczajowe nazewnictwo związków posiadających grupę karbonylową. W myśl owego nazewnictwa atom węgla sąsiedni do grupy karbonylowej nazywa się węglem α, kolejny bardziej odległy atom węgla nosi nazwę węgla β, dalsze litery alfabetu greckiego są ewentualnie przyporządkowywane kolejnym atomom węgla. A zatem według nazewnictwa zwyczajowego, wszystkie aminokwasy białkowe są α-aminokwasami o względnej konfiguracji centrum chiralności L (jeśli posiadają centrum chiralności) na atomie węgla α. Warto dodać, że glicyna jest achiralnym aminokwasem, ponieważ posiada płaszczyznę symetrii.
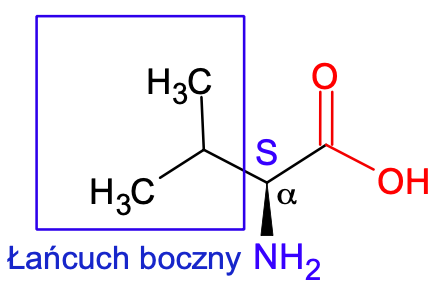
Ryc. 5. Struktura aminokwasów białkowych na przykładzie L-waliny
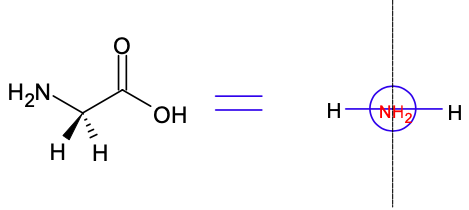
Ryc. 6. Cząsteczka glicyny i rysunek z perspektywy od grupy aminowej z widoczną płaszczyzną symetrii. Pogrubiony klin oznacza wiązanie skierowane do odbiorcy (przed ekran), a klin przerywany oznacza wiązanie skierowane od odbiorcy (za ekran).