Adrian Macion
Zakład Genetyki Bakterii, Instytut Mikrobiologii, Wydział Biologii UW
Pierwotniaki (Protista) są jednokomórkowymi organizmami jądrowymi (Eucaryota). W ogólnym modelu budowy cechy pierwotniaków są zbliżone do cech pozostałych komórek jądrowych. Podobnie jak grzyby czy rośliny posiadają one jądro komórkowe (możliwe są jednak specyficzne zmiany w obrębie liczby i architektury jądra – porównaj: orzęski). Procesy podziału i rozlokowania chromosomów do komórek potomnych przebiega z udziałem wrzeciona kariokinetycznego (o klasycznej budowie mikrotubuli). Ośrodkami formowania wrzeciona są bieguny wrzeciona podziałowego, które (w przeciwieństwie do komórek zwierzęcych) nie zawsze zawierają centriole. Protista stanowią niezwykle różnorodną grupę organizmów jednokomórkowych – zarówno pod względem struktury komórki jak i jej fizjologii, ekologii i możliwości formowania nawet bardzo złożonych kolonii. Kolonie możemy podzielić na złożone z osobników o identycznej strukturze i fizjologii (kolonie izomorficzne) lub złożone z osobników odmiennych (kolonie heteromorficzne). Kolonie heteromorficzne stanowią najwyższy stopień złożoności struktury i funkcji zespołów pierwotniaków. Pojedyncze osobniki mogą ulegać specjalizacji do pełnienia określonych funkcji. Specjalizacja ta w niektórych przypadkach jest tak daleka, że osobnik taki nie jest w stanie funkcjonować jako komórka samodzielna. Specjalizacja jest możliwa dzięki obecności różnorodnych organelli np. aparat gębowy z cytostomem, wodniczki tętniące, organelle obronne.
Pochodzenie
Nie które organelle (jądro, retikulum endoplazmatyczne, lizosomy, aparat Golgiego) powstały poprzez izolację kompartmentów otoczonych błoną w obrębie pojedynczej komórki. Zupełnie odmienną drogą ewolucyjną zaszło wykształcenie organelli częściowo autonomicznych (autonomia ta jest zachowywana poprzez obecność materiału genetycznego i częściowo niezależne systemy replikacji, transkrypcji i translacji). Teoria endosymbiozy zakłada, że eukariotyczna komórka powstała na drodze wchłonięcia komórki/komórek bakteryjnych i wzajemnej ewolucji tak powstałego układu. Symbiotyczne sinice miałyby przekształcić się w chloroplasty, a proteobakterie – w mitochondria. W trakcie tych przemian powstała również otoczka jądrowa, która wyodrębniła materiał genetyczny gospodarza w postaci niezależnego kompartmentu.
Struktura i fizjologia
Obecne u Protista systemy błon wewnętrznych i kompartmenty mogą być uorganizowane charakterystycznie dla danych grup pierwotniaków i odmiennie niż u komórek innych eukariotów np. wielokomórkowych. Skład biochemiczny i ultrastruktura tak podstawowych elementów jak błona biologiczna jest jednak typowy dla komórek eukariotycznych (dwuwarstwa lipidowa z białkami – model płynnej mozaiki usztywnionej cholesterolem). Na powierzchni błony otaczającej komórkę obecne są liczne glikoproteiny i glikolipidy (glikokaliks). U niektórych grup (patrz: nagie ameby) jest on silnie rozwinięty i tworzy osłonkę ochronną dla komórki. Większość pierwotniaków ma rozwinięty cytoszkielet, w którego skład wchodzą mikrotubule, mikrofilamenty i filamenty pośrednie. Rozwój cytoszkieletu charakteryzuje się dwoma odmiennymi modelami.
W cytoszkielecie dynamicznym rozwinięciu ulegają głównie mikrofilamenty aktynowe i białka motoryczne z nimi powiązane. Najbardziej widoczne jest to u ameb, których komórkę możemy podzielić na strefę ektoplazmy (rzadsza część cytoplazmy, która zawiera głównie elementy cytoszkieletu i pozbawiona jest organelli – mogłyby one zostać uszkodzone w procesie drastycznej zmiany kształtu ciała ameby związanej z ruchem komórki i fagocytozą) i strefę endoplazmy (gęsta optycznie cytoplazma o kształcie sferycznym i centralnej lokalizacji w komórce, zawiera kluczowe organelle takie jak jądro i systemy błon wewnątrzkomórkowych). Cytoszkielet statyczny zbudowany jest głównie z białek filamentów pośrednich i mikrotubuli. Przy wystarczającej gęstości tworzy on sztywną sieć podbłonową (korteks), która uniemożliwia zmianę kształtu komórki i fagocytozę. Dodatkowo korteks może być wzmacniany systemem blaszek i łusek, których większość powstaje przy udziale aparatu Golgiego. Komórki z rozwiniętym korteksem wykształciły specjalne miejsce w błonie, w którym dochodzi do fagocytozy i pinocytozy – cytostom (odżywianie), oraz miejsce związane z egzocytozą – cytopyge (wydalanie). Jest to szczególnie widoczne u orzęsków i wiciowców.
Większość pierwotniaków może poruszać się zarówno po podłożu stałym (forma pełzająca) jak i w toni wodnej (forma planktoniczna). Zmiana sposobu ruchu nie musi być ściśle związana ze zmianą morfologii komórki, jednak u pierwotniaków wyspecjalizowanych zmiana mechanizmu ruchu jest niemożliwa. Widzimy więc, że sposób ruchu komórki zależy od jej budowy. Różnorodność ogólnych kształtów i planów architektonicznych Protista jest ogromna, lecz można ująć ją w pary głównych typach:
- Wiciowce – dzięki grubej i sztywnej osłonce komórkowej (korteks, płytki białkowe i glikokaliks) kształt pierwotniaka jest w zasadzie niezmienny, a ruch warunkowany jest obecnością jednej lub kilku wici.
- Pełzaki – zmienny kształt ciała warunkowany rozwiniętym systemem włókien aktynowych umożliwia wytwarzanie pseudopodiów (wyróżniamy trzy zasadnicze rodzaje pseudopodiów: lobopodia – charakterystyczne dla ameb, płaskie i szerokie; filopodia – cienkie i długie włókna; retikulopodia – filopodia połączone mostkami cytoplazmatycznymi/anastomozami, całość systemu przypomina sieć).
- Słonecznice – ciało kuliste wykształcające delikatne filopodia.
- Sporowce – pozbawione organelli umożliwiających ruch.
- Orzęski – jedne z najbardziej charakterystycznych przedstawicieli Protista o ciele pokrytym gęstym systemem rzęsek, których ciałka bazalne połączone są siecią białek odpowiedzialnych za kontrolę regularnego ruchu rzęsek (kinetodesma). Pod błoną mogą znajdować się również pęcherzyki (alveole) i organelle obronny (trichocysty).
Pod względem strategii odżywiania się pierwotniaki dzielimy na dwie grupy: autotroficzne i heterotroficzne. Autotroficzny styl życia jest bardziej zaawansowany ewolucyjnie i jego powstanie wiąże się z endosymbiotycznym uzyskaniem odpowiednich organelli. Autotrofia może też być warunkowana poprzez czasową i fakultatywną obecność endosymbionta w ciele pierwotniaka. Ponadto wyróżnić można jeszcze charakterystyczna dla eugleny miksotrofię – która oznacza możliwość zmiany między strategią autotroficzną i heterotroficzną w zależności od warunków środowiska. W przypadku niedoboru światła i obecności w otoczeniu materii organicznej euglena wykazuje cechy heterotrofa, a warunkach odwrotnych z uwagi na obecność plastydów jest autotrofem.
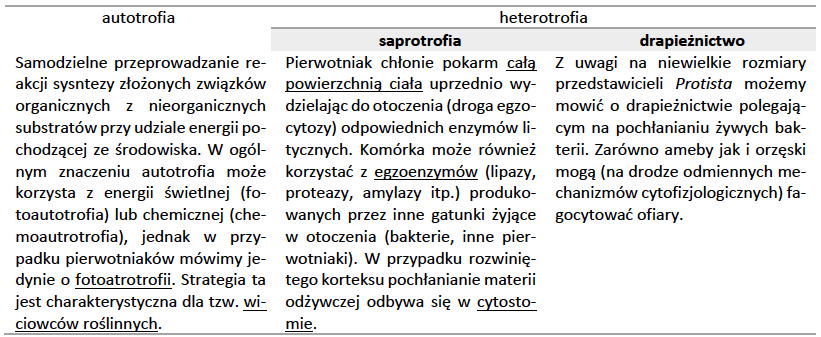
Pożywienie wprowadzone do wnętrza komórki na drodze fagocytozy lub pinocytozy zostaje otoczone błoną – powstaje wodniczka (wakuola) trawiąca, do której wprowadzane są enzymy lityczne np. proteazy, lipazy, esterazy, amylazy i nukleazy. Rozkład materiału pochłoniętego doprowadza do powstania monomerów, które są transportowane do cytoplazmy. Niestrawione resztki są usuwane przez cytopyge.
Z uwagi na wodne środowisko życia pierwotniaków organizmy te wyspecjalizowały się w usuwaniu nadmiaru wody z komórki przy udziale specjalnych organelli – wodniczek tętniących. W przypadku umieszczenia komórki w środowisku hipotonicznych (o stężeniu osmolitów mniejszym niż ich stężenie w komórce) woda intensywnie napływa przez pory w błonie i może doprowadzić do jej pęknięcia. Wodniczki tętniące usuwają ten nadmiar wody poprzez pobieranie jej z cytoplazmy i mechaniczny skurcz, który wyrzuca wodę poza komórkę w miejscu połączenia błony wodniczki z błona komórkową.
Pierwotniaki mogą dzielić się w sposób płciowy i bezpłciowy. W procesie bezpłciowym ciało pierwotniaka ulega podziałowi mitotycznemu na dwie identyczne komórki potomne. W przypadku rozmnażania płciowego dochodzi do wytworzenia gamet, które zlewając się ze sobą (cytogamia) doprowadzają do połączenia jąder komórkowych (kariogamia), wymieszania się materiału genetycznego i utworzenia zygoty. Gamety mogą być morfologicznie identyczne (określamy je wtedy mianem izogamet) lub różne (anizogamety).
Pierwotniaki wolnożyjące funkcjonują w wysoce zmiennym środowisku. Wykształciły więc one zdolność do tworzenia form przetrwalnych np. cyst. Cysty są to formy pierwotniaka, który spowalnia swoje reakcje metaboliczne do absolutnego minimum (które jest niezbędne do przetrwania) i otoczył komórkę organiczna barierą. Pancerzyk ten może być zbudowany z białek, glikoprotein śluzowych czy cukrów. Czasami ulega też inkrustacji i adkrustacji związkami nieorganicznymi (węglan wapnia, krzemiany i sole metali przejściowych), które nadają tym strukturom charakterystyczne barwy.
Systematyka
Systematyka Protista jest dziedziną wyjątkowo dynamiczną. Z uwagi na postępy w technikach badań genetycznych i coraz tańsze sekwencjonowanie genomów często dokonywane są reorganizacje ustalonego porządku systematycznego. Poniżej przedstawiono jeden z głównych, lecz nie jedyny prawidłowy model podziału Protista.
Wiciowce
Wiciowce są grupą bardzo liczna i różnorodną, zarówno pod względem morfologii jak i fizjologii komórek. Wśród nich możemy wyróżnić formy bardzo proste jak i niezwykle wyspecjalizowane pasożyty. Tylko dwie cechy są wspólne dla wszystkich wiciowców: organellum ruchu w postaci jednej lub kilku wici (o klasycznej budowie utrastrukturalnej) i rozmnażanie bezpłciowe na drodze podziału podłużnego (według długiej osi ciała). Wiciowce stanowią jedyna grupę pierwotniaków, w której prócz typowych form heterotroficznych możemy jeszcze wyróżnić roślinopodobne autotrofy:
1. Wiciowce roślinne stanowią grupę bardzo niejednolitą. Aparaty fotosyntetyczne wielu przedstawicieli oprócz chlorofilu zawierają również liczne barwniki dodatkowe, które nadają pierwotniakowi charakterystyczna barwę (żółtą, złocistą lub pomarańczową). Pewne gatunki w specyficznych warunkach mogą tworzyć zakwity, w trakcie których populacja wiciowców drastycznie rośnie. Zjawisko to nie jest obojętne dla innych organizmów w środowisku życia wiciowca. Produkty przemiany materii wydalane przez komórki gromadzą się w wodzie prowadząc do zahamowania wzrostu glonów, a ostatecznie nawet do śmierci samych wiciowców. Zakwit ulega wtedy samoograniczeniu i ostatecznie zanikowi. Dodatkowo zwielokrotniona populacja wiciowców zmniejsza natlenienie zbiornika na skutek intensywnego oddychania w nocy – może to prowadzić nawet do masowego śnięcia ryb.
a) Złotowiciowce są grupą licznie reprezentowaną w wodach słodkich i charakteryzują się bogactwem form. Wyróżniamy wśród nich formy planktoniczne, osiadłe, osłonkowe lub kolonijne. Dzięki dodatkowemu barwnikowi (chryzochromowi) chromatofory złotowiciowców mają barwę od żółtej po jasnobrązową. Materiałem zapasowym jest leukozyna, a cysty obudowują się ścianą wysyconą krzemionką. Formy kolonijne mogą przyjmować kształt drzewa, w którym każdy osobnik otoczony jest osłonką i przymocowany do dna lub innego osobnika stylikiem o właściwościach kurczliwych.
b) Bruzdnice cechuje bardzo charakterystyczna budowa. Ich ciało osłonięte jest sztywnym pancerzykiem z celulozy (często misternie żłobionym i asymetrycznym) i przedzielone dwiema bruzdami: bruzda poprzeczna otacza całą komórkę w pozycji równikowej dzieląc ją na część górną (episom) i dolną (hyposom); bruzda podłużna jest znacznie krótsza i ogranicza się do jednej strony ciała. W obu bruzdach znajdują się wici, których kombinacja ruchów umożliwia przemieszczanie się organizmu. Drugą charakterystyczną cechą bruzdnic jest budowa jądra, które w interfazie zawiera równoległe rzędy ziaren odpowiadających poszczególnym chromosom. Taki model budowy nosi nazwę dinokarionu i jest charakterystyczny wyłącznie dla bruzdnic. Bruzdnice z uwagi na obecność dodatkowych barwników np. dinoksantyny przyjmują barwę od zielonej po brunatną. Kolor komórki zależy też od obecności soli metali przejściowych w osłonkach. Glukoza powstała na drodze fotosyntezy jest gromadzona w formie skrobi.
c) Eugleny mają klasyczną morfologię wiciowca. Prócz ich ciała zagłębia się w formie ampułki, z której wystają zazwyczaj dwie wici o różnej długości. Wić dłuższa związana jest z lokomocją, a krótsza odpowiada za „naganianie” pokarmu do wnętrz ampułki, której ściany nie zawierają sztywnej pellikuli i umożliwiają pochłanianie pokarmu. Są to jedyne wiciowce, których materiałem zapasowym jest paramylon (polisacharyd złożony z reszt glukozy połączonych wiązaniem β 1-3 glikozydowym, w jego syntezie uczestniczą zarówno chloroplasty jak i mitochondria). Pellikula niektórych gatunków charakteryzuje się sprężystością i umożliwia to zmianę kształtu ciała (metabolia). Osłonka ta może mieć budowę nieciągłą i być poprzebijana otworami, przez które wiciowiec wydziela śluz. W przedniej części komórki znajduje się stigma – aparat detekcji światła.
2. Wiciowce zwierzęce posiadają bardzo cienką (a co za tym idzie giętką) pellikule i pozbawione są aparatu fotosyntetycznego. W grupie tej wyraźnie widać ewolucyjną tendencję do pasożytniczego trybu życia.
a) Wiciowce kołnierzykowe (Choanoflagellata) są wiciowcami wolnożyjącymi, których ciało w przedniej części otoczone jest delikatnym kołnierzykiem cytoplazmatycznym, w którego centrum znajduje się wić. Kołnierz ten pełni istotną rolę w procesie odżywiania się pierwotniaka. Niestrawione resztki usuwane są drogą egzocytozy u podstawy wici. Formy osiadłe wytwarzają stylik, którym wiciowiec przytwierdza się do podłoża.
b) Świdrowce są wiciowcami wyłącznie pasożytniczymi o ciele wydłużonym, często wstęgowatym z jedną wicią skierowaną ku przodowi ciała. Wić ta często zagina się wzdłuż krawędzi ciała ku tyłowi i tworzy błonę falującą. Z uwagi na pasożytniczy tryb życia i obecność więcej niż jednego żywiciela w cyklu rozwojowym świdrowce przystosowały się do bytowania w różnych środowiskach. Jednym z mechanizmów tego przystosowania jest występowanie wiciowca w kilku różnych formach – polimorfizm:
- trypomastigota – wrzecionowata, z jądrem i kinetoplastem w pobliżu bieguna tylnego, wić i błonka falująca przebiegają wzdłuż całej komórki kończąc się na jej przednim biegunie. Postać ta spotykana jest we krwi obwodowej kręgowców;
- epimastigota – forma wrzecionowatego kształtu z wicią, błoną falującą, kinetoplastydem i jądrem umiejscowionymi od strony przedniego bieguna komórki; jest to postać spotykana we krwi infekowanego organizmu;
- promastigota – forma wrzecionowata, kinetoplast, kinetosom i wić położone w pobliżu przedniego bieguna komórki, przy czym brak jest błony falującej; postać spotykana w trakcie bytowania w ciele bezkręgowców;
- sferomastigota – forma sferyczna, jądro i kinetoplast położone centralnie, występuje wić, jednak zwykle brak błony falującej.
- amastigota – forma sferyczna, jądro i kinetoplast położone centralnie, brak wici i błony falującej – forma nieruchliwa.
c) Parabasalia są cudzożywnymi protistami, u których mitochondria uległy przekształceniu w hydrogenosomy – organelle, w których dochodzi do wytwarzania ATP w procesach beztlenowych związanych z wydzieleniem wodoru. Niektóre gatunki współżyją z termitami zasiedlając ich jelito i przy udziale bakterii symbiotycznych umożliwiają trawienie celulozy. Wici ułożone są w skupisku na przednim biegunie ciała, jedna wić może zakręcać ku tyłowi i tworzyć błonę falującą. Przez środek ciała przebiega aksostyl – sztywny pęk mikrotubul.
Korzenionóżki
Odznaczają się brakiem wyodrębnionego aparatu ruchu. Dzięki ameboidalnemu kształtowi ciała ruch odbywa się w sposób pełzakowaty. Cytoplazma jest dynamiczna, można to zauważyć obserwując ruch ziarnistości w obrębie komórki. Rozmnażanie następuje poprzez prosty podział poprzeczny podobny do mitozy, a w niektórych grupach występuje również rozmnażanie płciowe.
- Ameby – pomimo asymetryczności ciała ameby możemy jednak w niektórych przypadkach określić pewną biegunowość. Przejawia się ona w wytwarzaniu na końcu ciała collopodium – lepkie pseudopodium, które przytwierdza się do podłoża, a po odrywaniu zabiera ze sobą cząstki pokarmowe. Część ameb wytwarza osłonki, które nie są przepuszczalne dla pseudopodiów. Osłonki te cechują się dużą różnorodnością form i materiałów, z których są zbudowane. Mogą być inkrustowane i adkrustowane związkami nieorganicznymi (w tym barwnymi solami metali).
- Otwornice są grupą prawie wyłącznie morskich organizmów. Wszystkie mają komórki osłonięte skorupą wzmocnioną węglanem wapnia, która poprzeszywana jest licznymi otworami umożliwiającymi wysuwanie retikulopodiów/filopodiów.
Promienionóżki
Do grupy tej należą pierwotniaki wyłącznie słodkowodne o okrągłym ciele z wystającymi filopodiami. Niektóre gatunki mają szkielet zbudowany z igieł krzemionkowych. U większości słonecznic w centrum endoplazmy znajduje się centroplast – pęcherzyk z zamkniętym ziarnem załamującym światło. Prawdopodobnie pełni funkcję ośrodka ruchu, ponieważ do jego błony dołączone są podstawy aksopodiów.
Sporowce
Do sporowców należą wyłącznie protisty pasożytnicze, które w stadium dojrzałym nie mają żadnych organelli ruchu. Cykl życiowy zaczyna się od ruchliwej formy – sporozoitu. W ciele żywiciela sporozoit przekształca się w trofozoit, który przechodzi w stadium schizonta. Jądro schizonta dzieli się intensywnie, a cała komórka rozpada na schizogonia. Następnie dochodzi do wytworzenia anizogamet i zapłodnienia. Pierwszy podział zygoty jest mejotyczny i jego wyniku powstają ruchliwe sporozoity.
Orzęski
Orzęski stanowią grupę najbardziej zwartą wśród wszystkich Protista. Wyróżniają się one na tle innych pierwotniaków kilkoma charakterystycznymi cechami. Ich organellami ruchu są rzędy rzęsek, które u form pierwotnych pokrywają całe ciało, a u bardziej wyspecjalizowanych ulegają częściowej redukcji. U niektórych form rzęski zlewają się w sztywne cirri, które ulokowane są po brzusznej stronie ciała i uczestniczą w lokomocji. Otwór gębowy orzęsków nazywany jest cytostomem, w obrębie którego dochodzi do fagocytozy i pinocytozy. Jądra komórkowe orzęsków charakteryzują się dymorfizmem: wyróżniamy mikronukleus biorący udział w procesach płciowych i makronukleus, który dzieli się amitotycznie w trakcie podziału komórki. Zjawisko rozmnażania płciowego u orzęsków określane jest mianem koniugacji.
Podstawy parazytologii Protista
Pasożytnictwo jest jednym z antagonistycznych oddziaływań między organizmami, które polega na czerpaniu korzyści przez jeden organizm kosztem drugiego. Pasożyt żerując na żywicielu (który zazwyczaj jest dużo większy od pasożyta) osłabia jego organizm, jednak w zdecydowanej większości przypadków docelowo nie prowadzi do śmierci. Do zalet pasożytniczego trybu życia można zaliczyć: stabilność środowiska, ruchliwość żywiciela i dostępność pokarmu. Pasożyt stale musi chronić się przed mechanizmami obronnymi żywiciela i rozmnażać się w celu utrzymania ciągłości infekcji. W procesie ewolucji pasożyty przystosowały się do środowiska ciała żywiciela, a tym samym utraciły zdolność do życia w środowisku zewnętrznym. W związku z tym, że pasożyt przez zdecydowaną większość swojego czasu życia pozostaje związany z żywicielem możemy wyróżnić kilka ich typów:
- Żywiciel ostateczny – organizm, w którym pasożyt odbywa końcowy etap swojego rozwoju i rozmnaża się płciowo.
- Żywiciel pośredni – organizm, z którym pasożyt jest związany w trakcie swojego cyklu życiowego, ale nie rozmnaża się w nim płciowo.
- Żywicielami przypadkowymi są gatunki, które zazwyczaj nie są żywicielami danego pasożyta, lecz w bardzo sprzyjających warunkach mogą się tymi pasożytami zarazić i nie ograniczają jego rozwoju.
- Żywiciel rezerwuarowy (parateniczny) nie jest niezbędny w cyklu życiowym pasożyta, pasożyt żyjąc w nim nie traci swej wirulentności, ale nie czyni też szkody temu organizmowi.
- Żywiciel główny – gatunek w którym, w określonych warunkach ekologicznych pasożyt występuje w największej częstotliwości i intensywności.
- Żywiciel uboczny jest środowiskiem życia dla pasożyta tylko w pewnych warunkach i nie umożliwia pasożytowi pełen rozwój.
Podsumowanie:
Królestwo Protista jest najdziwniejszym ze wszystkich. Nie dość, że zostało utworzone sztucznie (w jego skład wchodzą organizmy wywodzące się od innych przodków, a cała grupa jest parafiletyczna), to zawiera jedne z najbardziej niesamowitych organizmów w znanej nam biologii. Dzięki temu jest ono niezwykle ciekawym zagadnieniem, w którym na każdym kroku czeka coś niezwykłego.